
0 просмотренных постов скрыто






Ответ на пост «Когда зашли в супермаркет»
Навеяло постом)
Кто помнит, во времена ковида был этакий онлайн-бунт в Москве.
К сожалению, поздно на него попала, большую часть перлов пропустила, но чуть заскринила.
Возможно, половцы и печенеги все ещё где-то там)
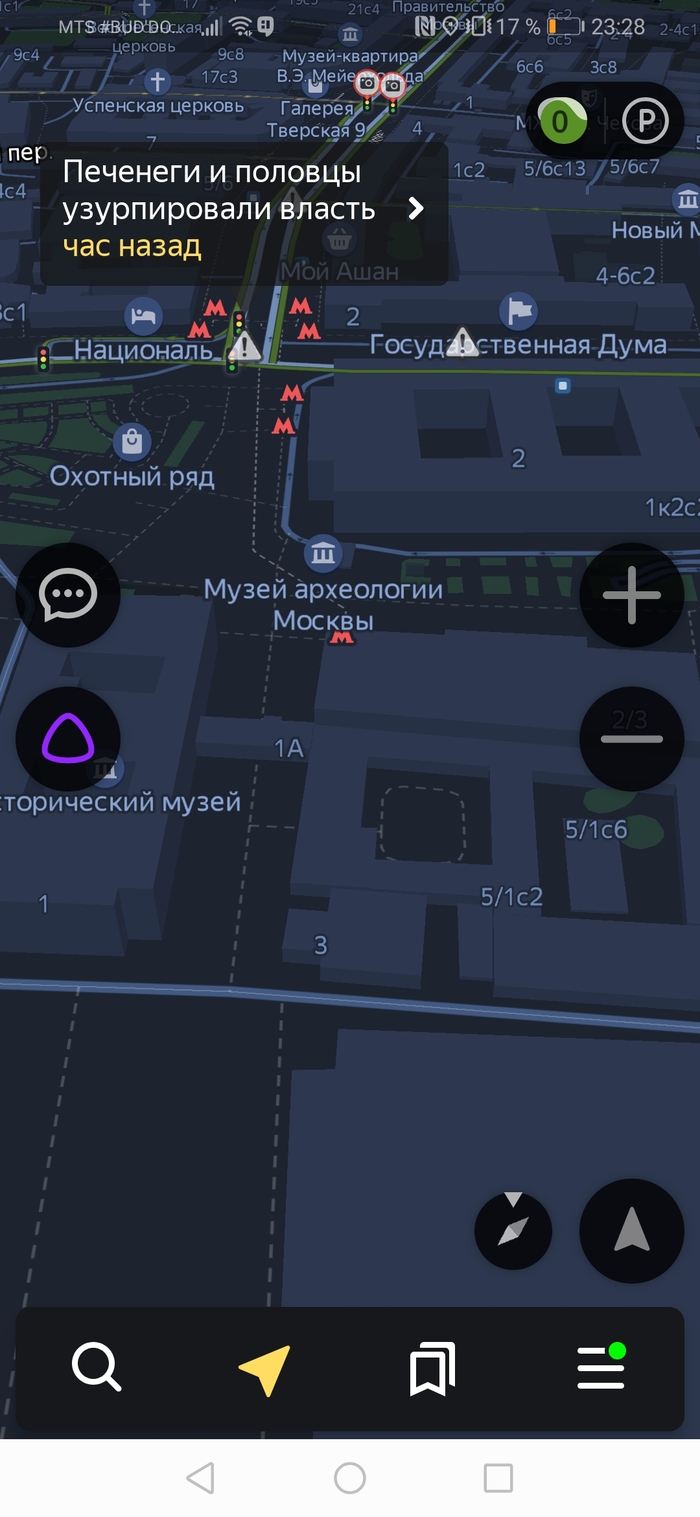
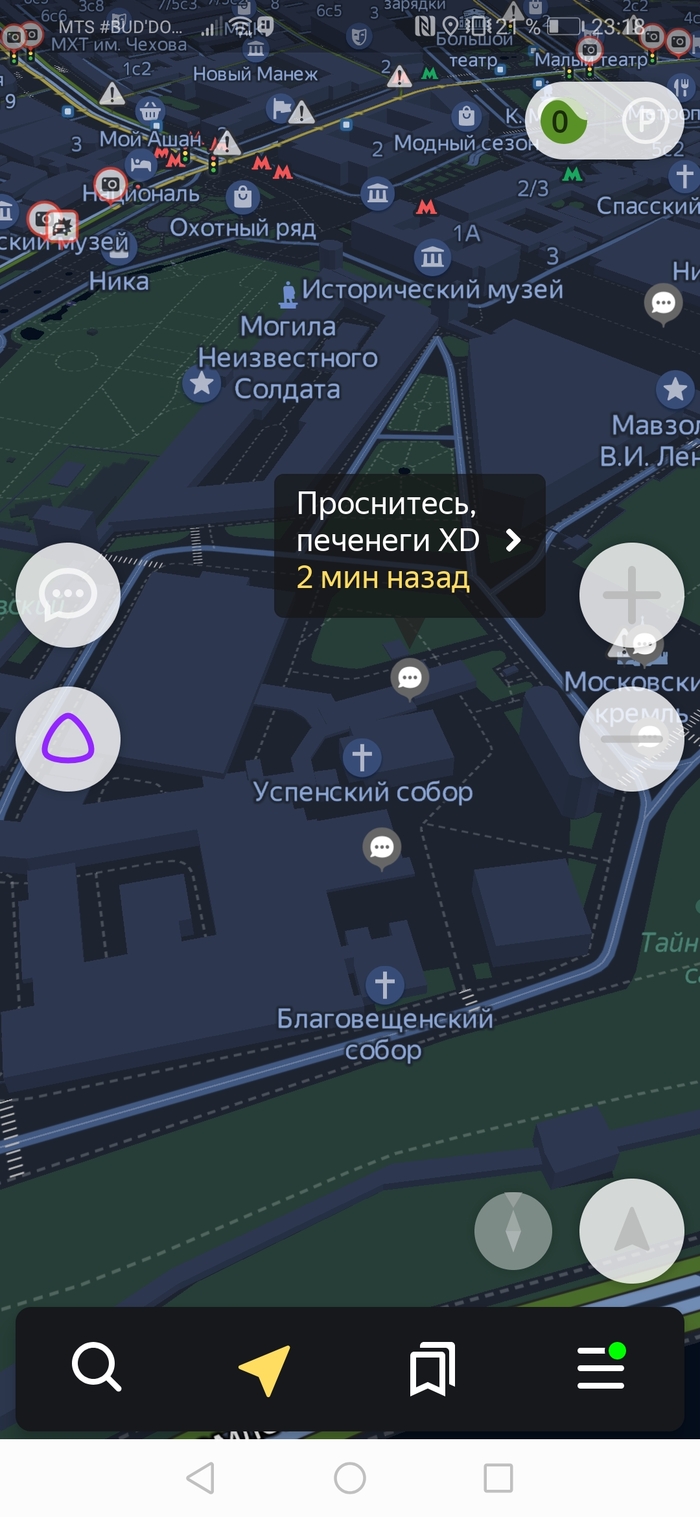
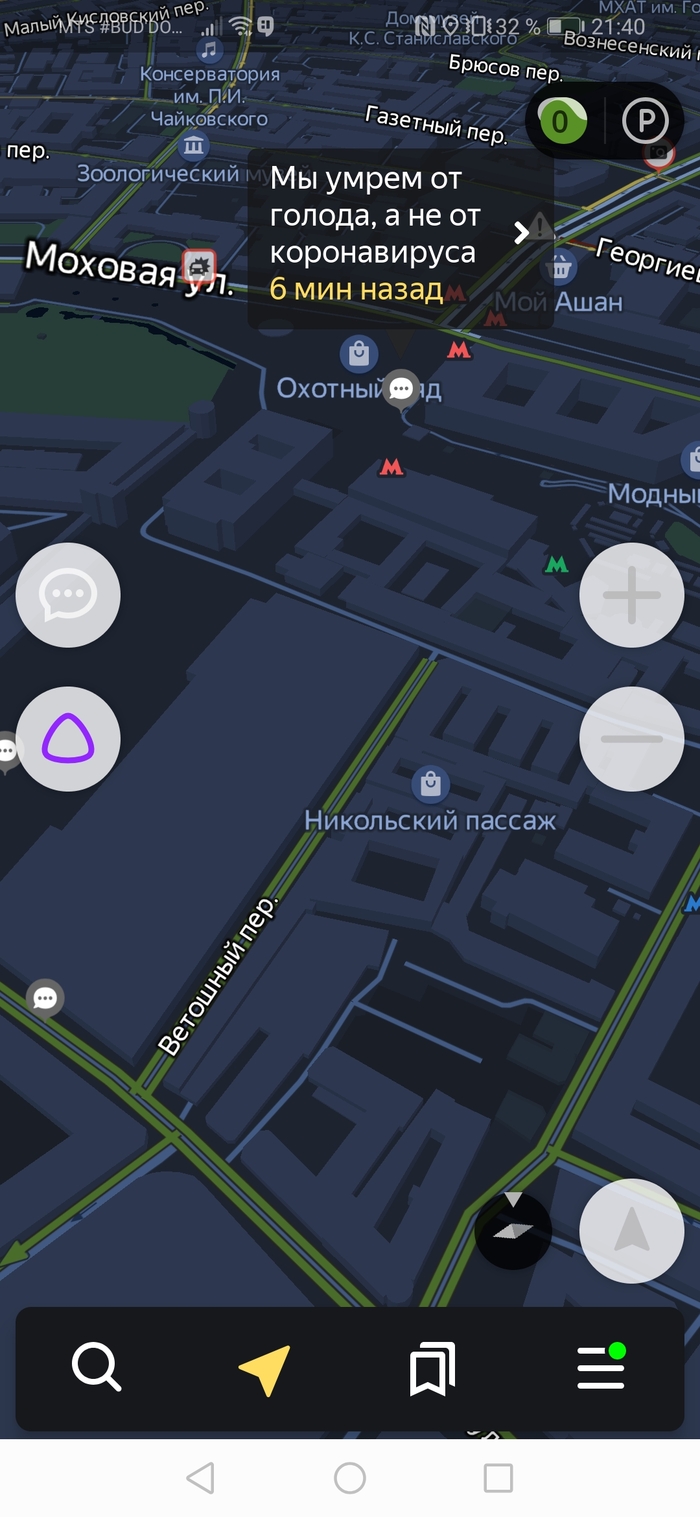
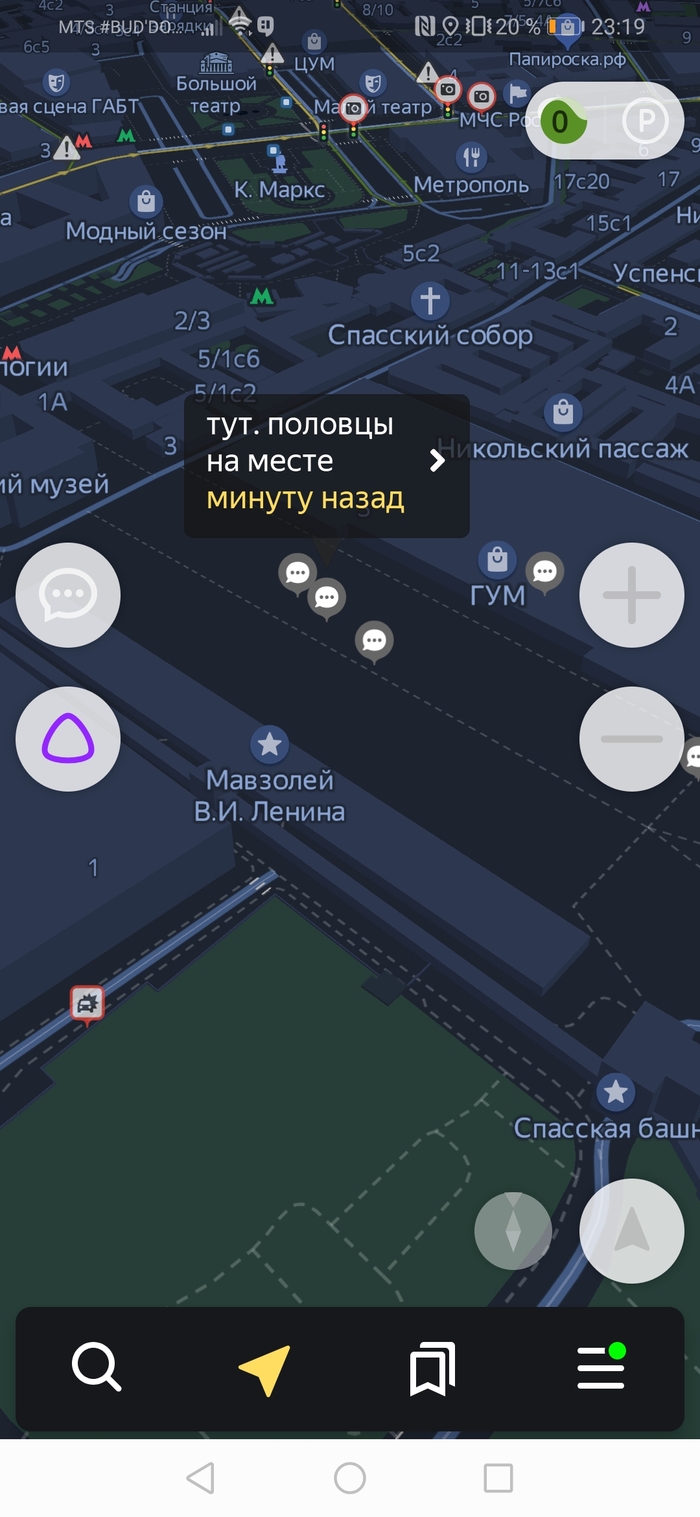

Скрины по отношению к власти прикладывать не буду, а то не дай боже тег политика влепят)
Показать полностью
5

Когда зашли в супермаркет
Прислали подписчики:)


Показать полностью
2

Ребята в теме

Показать полностью
1

Деполитизированная миграция протогерманцев R1b в Европу. Этимология слова "варяг"
Во времена "царя Гороха" на смену культу Рода пришел культ Яра. Произошло это событие в окрестностях острова Рюген на "море варяга", Балтийском море, в 5514 г. до н.э. (нулевой год Рода, ближе всего к этой дате Константинопольская (Девятеричная) 5509 г. до н.э., Десятеричная 5510 г. до н.э. и Одиннадцатиричная 5511 г. до н.э. эры) и известно как "сотворение мира", после чего в 5184-4954 гг. до н.э. была основана Аркона - столица Яровой Руси, позднее называемой греками "Европа". Население Яровой Руси так и называло себя верящими в Яра, в-Яра верящие, и, после аналогового словообразования (суффикс -аг по модели бродяга), в-Яра-г, варяг. Это пример фонетической адаптация при участии субпроцессов:
Редукция: Упрощение сочетания «въяр-» → «вар-» (как в «въезд» → «везти»).
Эпентеза: Вставка [г] в конце для соответствия модели -аг (ср. «юродивый» → устар. «юродга»).
Суффикс -аг в русском языке часто указывает на профессию, занятие или характеристику лица (архаичные примеры: бедняга, судага).
Из-за таяния ледников и глобального повышения уровня океана на 2–4 м уровень Балтики в те времена был на 4–7 м выше современного в районе Финского залива и до 15 м выше в южных районах. Южные районы (современные Германия, Польша) были затоплены, и были изрезаны лагунами и косами. Вследствие более высокой солености, Балтийское море было богато треской, камбалой, в нем обитали киты. По его берегам селились тюлени и образовывалось большое количество поселений человека. В течение последующих двух тысячелетий был пик голоценового климатического оптимума (7000–5000 лет до н.э.), с которым связана дегляциация Лаурентидского ледника в Северной Америке (около 6000 г. до н.э.).
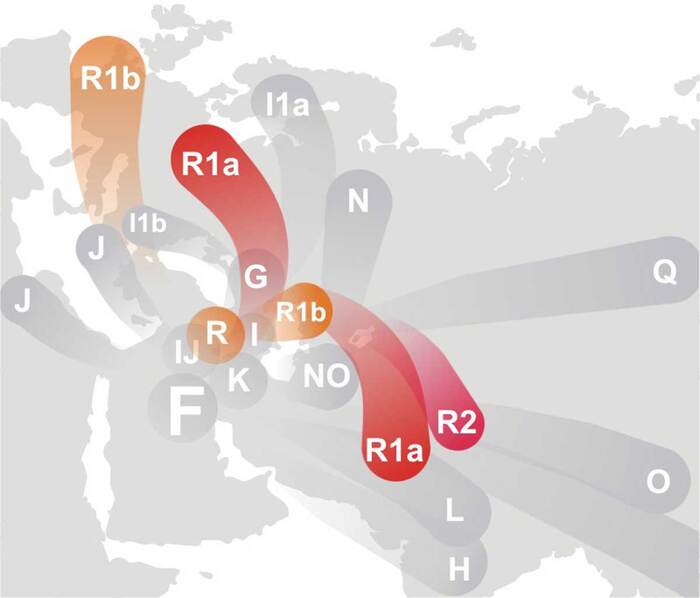
Основные пути миграции гаплогруппы R1.
Тогда на Балтике существовали следующие мезолитические культуры:
1. Эртебёлле (5400–3950 до н.э.)
Ареал: Южная Скандинавия (Дания, юг Швеции), север Германии.
Хозяйство: Морское рыболовство (треска, тюлени), сбор моллюсков, охота на оленя.
Артефакты: Керамика с остродонными сосудами, костяные гарпуны, микролиты.
2. Нарвская культура (5300–1750 до н.э.)
Ареал: Восточная Прибалтика (Эстония, Латвия, Литва).
Хозяйство: Охота (лось, бобр), речное рыболовство, использование янтаря.
Артефакты: Костяные наконечники, шлифованные топоры из сланца, керамика с ямочно-гребенчатым орнаментом.
3. Культура Кунда (8000–5000 до н.э.)
Ареал: Северо-восток Балтики (Финляндия, Карелия).
Хозяйство: Охота на тюленей, рыболовство, использование лодок-долблёнок.
Артефакты: Костяные рыболовные крючки, гарпуны с зубцами.
В Восточной Прибалтике в это время жили позднекундские / ранненарвские охотники-собиратели. Их остеологический материал изучён, не столь массово, однако десяток хорошо датированных и опубликованных геномов (могильник Звейниеки в Латвии, Донкалнис и Спигинас в Жематии, Кретуонас в Вост. Литве, отдельные скелеты с островов Эзель и Рухну в Эстонии) позволяет увидеть устойчивую картину Y-хромосомных линий, зафиксированные в выборке 5600–5400 BC:
I2a1b-M423 (позднее делится на L621/Dinaric) ≈ 25–30 % • линия балкано-карпатского происхождения (из Живиной Руси), занесённая на север, очевидно, ещё в эпоху раннего Дриаса. • сегодня её потомки — целое «островное море» Dinaric-South (I-Y3120) на Балканах и Dinaric-North (I-Y18331, I-Y4460, …) в Восточной Европе. Прибалтийские подкладки ближе всего к Y18331→CTS4002 / YP196.
R1a-M459* (xM417) ≈ 15–20 % • прародитель огромного «арийско-славянского» R1a-M417; • в течение III тыс. до н. э. из неё появляются ветви Z93 (азиатская) и Z283 (европейская); у балтов закрепляется Z283→Z280→CTS1211 / YP1361 / YP4758 и др.
Q1a2-M25 ≈ 5 % • арктико-сибирская линия, попавшая в Восточную Прибалтику ещё в позднемезолитическое время; • ныне даёт редкие прибалтийско-финские подветви Q-L939 и Q-YP1669.
Что стало с этими линиями к XXI в.:
1) R1a M459* → M417 → Z283 → Z280 → CTS1211 → – YP1361, YP605, YP237 & др. — «балтийский гребень» (до 35 % у латышей, 25 % у литовцев, 15 % у белорусов).
Итак, во времена "сотворения мира" на Балтику мигрировали R1a-M459* (иногда обозначают R1a* или R1a-“pre-M417”). Узловая мутация M417, из которой позже вырастут Z283, Z280, M458, появится существенно позже, в степном “котле” рубежа V–IV тыс. до н. э. и войдёт в Прибалтику лишь вместе с культурой шнуровой керамики (~2900–2500 гг. до н. э.).
Западнославянские (ободриты, вильцы, бодричи, лютичи) клады X-XI вв. изучены на могильниках «Fischerinsel», «Ralswiek», «Rostock Dierkow». Типичные Y-линии – R1a-M458, R1a-Z280, I2a-CTS10228, N1a-M278. R1b-U106 – единичные, < 5 %. «Wolinians» (о-в Волин, X–XI в.) – аналогичная картина.
2) I2a1b M423 → L621 (‘Dinaric’) → CTS10228 → Y3120 → – Y18331/CTS4002 (пик на бывших территориях Живиной Руси в Боснии 50 %, в Хорватии 35 %, до 9 % у литовцев, 7 % у белорусов).
Во времена "сотворения мира":
уже присутствует I2a1b-M423 (на нескольких индивидах из Звейниеки, Спигинаса, Донкалниса). • Линия L621 (“Dinaric”), а из неё и CTS10228, выделится позже (ориентировочно 4500–3500 гг. до н. э. в зоне Карпато-Балкан - Живиной Руси). ⇒ Прямого CTS10228 в VI тыс. до н. э. ещё нет, но его непосредственный предок – M423 – составляет заметную долю мужского населения (20-30 %).
R1b P297* → (часть ушла на юг) → M269 → L51 → P312/U106… — основа современного западно-европейского R1b; Лишь микроскопические «пережитки» древнего P297* фиксируются в Прибалтике, Финляндии и на Южном Урале.
По политическим мотивам и предвзятости археологических изысканий считается, что R1b-P297* (xM269) составляла около 5514 г. до н.э. порядка 45–50 % всей популяции. Так ныне проявляется немецкий национализм. По сегодняшним националистическим норманистическим представлениям считается, что R1b-P297* через 1–1,5 тыс. лет даёт разветвление P297→M269; её потомки сегодня — весь «западный» R1b (L51/P310 → P312, U106, …) и степной Z2103. То есть ложно утверждается, что немцы - коренные обитатели Европы. Хотя конкретные балтийские P297* в XX–XXI вв. почти исчезли; самые близкие «реликты» обнаруживаются у единичных латышей и карелов как R1b-YP257/R1b-YP142. И действительно, Прибалтика, ввиду своего финно-угорского происхождения населения, по-видимому, оказалась не в зоне политизированных археологических изысканий.
Q1a2 M25 → L939 → YP1669 — сегодня ≤ 1 % у эстонцев, ливов, коми-пермяков.
N1a (балты, финно-угры):
Первые N1a1a-M178 (не M278) появляются лишь в горизонтах культуры гребенчатой керамики / позднешнуровых комплексов Финляндии и Карелии (~2300–1800 гг. до н. э.). Мутация M278 относится к другой ветви (N1a2) и в Прибалтике не зафиксирована вовсе. Следовательно, никаких “предковых” N-линий во времена "сотворения мира" на Балтике ещё не было, то есть не было балтов вовсе.
Таким образом, «ядро» мужского генофонда восточной Балтики середины VI тыс. до н. э. составляли ветви I2a1b-M423 и R1a-M459., которые пережили бронзовый век и сегодня проявляются соответственно в балкано-балтийском I-CTS10228 и в балтийско-славянском R1a-CTS1211/YP1361. Древнейший известный субклад R1a-M459, обнаружен у мезолитического охотника из Южного Оленьего Острова (Карелия, 5500–5000 до н.э.), вероятно, присутствовал у популяций Восточной Балтики.
Ниже – «скелетная» схема того, как пра-R1b-P297 из восточноевропейских лесостепей превратилась в две основные сегодняшние ветви R1b-L51 (западная Европа) и R1b-Z2103 (южное Причерноморье, Кавказ, Балканы).
I. Откуда берётся P297:
17 000–12 000 ВС (BC - система отсчёта, используемая в радиоуглеродном датировании, где 1950 год принят за начальный год отсчёта) (поздний ледник / Allerød) • Восточная Сибирь и Алтай – археогенетические носители «R*» (MA-1, Afontova-Gora). • Часть этой популяции движется на запад и, смешавшись с европейскими охотниками, создаёт т. н. EHG («Eastern Hunter-Gatherers», Восточноевропейские охотники-собиратели).
≈ 11 000–8 000 ВС (мезолит) • Волжско-уральские, карельские, лапоноидные стоянки – ровесники получают Y-ДНК R1b-P297* (Karelia_HG, Samara_HG). • Климат теплеет → EHG расселяются от Урала до Балтики. • Само P297 в Европе – уже европейское, а не сибирское явление.
II. «Балтийская» фаза P297*
≈ 6500–5500 ВС • Позднемезолитические могильники Звейниеки (Латвия), Спигинас, Донкалнис (Литва) дают серию скелетов R1b-P297* (без М269). • Распространение ограничено Северо-Восточной Европой; к западу от Вислы P297 почти нет.
III. Рождение M269 и первых степных субклассов
Хвалынск (средняя Волга) 5200–4500 ВС – cкелет I0122: R1b-M269*; – здесь же L23xZ2103 и др. базальные линии.
Ранний Repin / Ямная-I 4400–3800 ВС – на нижней Волге фиксируются уже R1b-L23.
→ Отныне P297* разделилась географически: • северный «остаток» остаётся в Прибалтике (и постепенно вымирает); • южная линия M269 начинает масштабную экспансию со степи.
IV. Два «крыла» M269
R1b-Z2103 (юго-восточное крыло) 3300–2600 ВС – классическая Ямная (Самара, Саратов, Украина) практически поголовно Z2103. 3000–2400 ВС – катакомбная культ., MBA Кавказа, ранние балканские курганы – всё ещё Z2103. Позднее: армяне, северо-кавказцы, болгары, греки (до 30 % в ряде горных районов).
В VI-VIII веках в Прикаспия (нижняя Волга, Прикубанье, Северный Казахстан) и на соседнем Северном Кавказе фиксируются первые R1b-L23* (xZ2103, xL51) – это реальная предковая стадия для европейских эрбинов: L51, P312, U106.
На самом Кавказе (предгорный и внутригорный) есть несколько древних и множество современных носителей R1b-L23*; дальше по иерархии (L51 и её субклады) до сих пор не встречены. То есть эрбины пришли в прегорья Кавказа из Прикаспия.
R1b-L51/P310 (западное крыло)a) «Балкано-карпатский» инкубатор Считается, что появились в 3800–3000 ВС (фактически лишь в VIII-X веках) – в ямных группах Болгарии, Румынии, Венгрии появляются первые L51*, L52*, ещё без P312/U106.b) 2900–2500 ВС – культура колокотообразных кубков (Bell Beaker) – старейшие чистые L51→P312 датированы 2750–2600 ВС (фактически лишь в VIII-X веках) (Карпаты, Моравия); – вместе с «кубками» линия взрывообразно идёт: Рейн → Атлантику → Британские о-ва, а по ветке U106 – в Северную Германию и Скандинавию.c) 2500–1500 ВС (фактически лишь в VIII-X веках) – P312 подразделяется на DF27 (Иберия), L21 (Британия-Ирландия), U152 (Центр. Европа, Альпы); – U106 становится «германской» доминантой.
В Позднеантичном малом ледниковом периоде (536–660 гг. н.э.) произошла серия мощных вулканических извержений (536, 540, 547 гг.), выбросивших сульфатные аэрозоли в атмосферу. Это привело к глобальному похолоданию на 1.5–2.5°C и сокращению осадков. Далее из-за таяния ледников Тянь-Шаня и Памира произошло снижение стока Трансаксонских рек — двух крупнейших водные артерий Средней Азии: Сырдарьи и Амударьи, которые впадают в Аральское море на территории Казахстана.
Из-за учащение засух и пылевых бурь в Прикаспии, опустынивания земель огузы и саки мигрировали из их исконных земель в Трансаксонии (регион Тянь-Шаня и Памиро-Алая). Хотя считается, что огузы и саки мигрировали в сторону Европы в две разные волны:
• Саки — ираноязычный народ, входивший в скифо-сакский культурный ареал (восточноиранская группа). • Огузы — тюркоязычные племена, чьи потомки говорят на огузских языках (азербайджанский, турецкий, туркменский).
Хронологический разрыв • Саки упоминаются в источниках с VIII в. до н.э. до раннего Средневековья. • Огузы формируются как отдельная группа к IX–X вв. н.э., спустя столетия после ассимиляции саков.
На самом деле они мигрировали в одно и то же время, археологический сдвиг для них равен римскому сдвигу в 1200 лет. Это сдвиг и является причиной, по которой останки саков не типируют, относя их в глубокую древность и не находя возможности откопать их останки. Саки и огузы - это и есть те самые гунны-печенеги, а потом и кипчаки-половцы, которые сформировали европейское население.
Историко-лингвистический парададокс заключается в том, что пратюркский язык реконструируется в XIII–VIII вв. до н. э. (отражён в изоглоссах с монгольским, тунгусским, яньцай/аланьскими экзонимами), письменные признаёт к V в. н. э. Прагерманский (в том числе фонологические сдвиги *p > f, *t > þ и т.п.) считается уже завершённым к 500 г. до н. э. и задокументированным латинскими и греческими транскрипциями имён в I в. до н. э. Чтобы «огузский» язык стал прагерманским, нужно перенести его на 1200 лет назад, а германский – на столько же вперёд. Этот сдвиг разрушает всю известную палеолингвистическую картину Евразии (индоиранские, кельтские, италийские, балто-славянские изоглоссы) и отвергается современной наукой.
Чтобы постулировать указанный свдиг на 1200 лет, нужно: • постулировать глобальный сбой ^14С и всех калибровочных независимых шкал на ±2500 лет; • отменить археологическую преемственность культур в Европе и Центральной Азии; • переписать сравнительно-историческую грамматику индоевропейских и алтайских языков.
Тем не менее, из ≈10 000 секвенированных древних Y-хромосом «коллекция» U106/P312 все же обнаружена на востоке во времена эллинизма (с учетом сдвига в 1200 лет это те же VI - VIII века) через военные, торговые и колонизационные контакты.
Огузы и саки делят между собой «восточно-степные» ветви R1b-M73 и R1b-Z2103 (PH155); – Саксы опираются на «западно-европейские» R1b-U106 и (частично) R1b-P312; – прямого родства между «огузо-сакской» и «саксонской» R1b-компонентами нет, их связь ограничивается общим предком L23 эпохи ранней Ямной культуры.
Однако все же существуют единичные находки U106/P312-комплекса на востоке от Германии (приведены далее), просто у националистически настроенной археологии нет указания искать останки с U106/P312-комплексом за Карпатами.
Теперь рассмотрим, куда на дереве R1b попадают упомянутые клады.

Древо субкладов R1b
Современные популяции, считающиеся прямыми наследниками раннесредневековых огузов (туркмены, огузские кланы каракалпаков, часть сельджуко-анатолийцев), показывают: • R1b-M73 ≈ 15–35 %, иногда в субкладах KP1/L1405; • R1b-Z2103 (в т. ч. PH155, L584) ≈ 10–25 %; • редкие R1b-L23* (xZ2103,xL51); • минимальные количества «европейских» U106/P312 – почти всегда результат более поздней кульминации (генофонд Балкан и Вост. Европы у турок, русские/украинские примеси у каракалпаков и т. п.).
Действительно, палеогенетика могильников Schleswig-Holstein, Нижней Саксонии, Восточной Англии подтверждает: • R1b-U106 (крупные ветки Z381→Z156→L48 и Z301) ≈ 50–60 % мужских скелетов; • R1b-P312 (DF27, U152, кельто-романское наследие) 8–15 %; • минимальные следы Z2103 (<1 %) и полное отсутствие M73. Современные саксонские районы Германии / Англии сохраняют ту же картину.
Считается, что предки европейцев (саки, аланы-ясы) принадлежат к иранской ветви индоевропейских народов. Саки (VIII в. до н.э. — IV в. н.э.) считаются предками алан (I–XIII вв.), а ясы (упомянуты в венгерских хрониках) — западной ветвью алан. Так, курганные погребения в стиле «звериного искусства» (Тянь-Шань, Причерноморье) демонстрируют преемственность сакской и аланской культур. Т-образные склепы в Крыму (IV–V вв.), характерные для алан, повторяют традиции сакской архитектуры.
Анализ ДНК из сакских курганов (Пазырыкская культура) и осетин выявил общую гаплогруппу R1a-Z93, характерную для индоиранских кочевников. Однако у ясов фиксируется примесь угорских маркеров от гуннов-мадьяр из Приуралья. Таким образом, ясы-аланы — не прямые потомки саков, а результат синтеза сакских, мадьярских и местных кавказских (осетинских) линий предков.
Действительно, Аммиан Марцеллин (IV в.) отождествлял алан с древними массагетами — племенем, родственным сакам. Венгерские хроники XIII в. называют ясов «асами», используя термин, восходящий к аланскому самоназванию «асы». Их столица была в городе Азов - Асгард.
Саков образно относят к степному «котлу». С середины V тыс. до н. э. (фактически, VI в. н.э.) из него выделяется M269, а затем L23.
Из Степи в саму Европу идут две волны: – ранняя ямная Z2103 через Балканы; – поздняя ямная/L51 через Карпаты, породившая культуру «колокотов» и ставшая прародительницей 80–90 % современного западно-европейского R1b.
У огузских тюрков (туркмены, огузские предки современных турок-огузов) и у древних сако-скифских культур Центральной Азии встречаются одни и те же «степные» ветви R1b – прежде всего R1b-M73 и R1b-Z2103 (часто в форме субклада PH155). R1b-M73 отделился от общего предка R1b ~16 000 лет назад и не связан с европейскими субкладами. R1b-Z2103 и R1b-U106 — параллельные ветви внутри R1b-M269, разделившиеся ~5 500–6 000 лет назад. Ни одна не является предковой для другой.
У саксов (германцев Северной Германии и Англии) доминируют «западноевропейские» ветви R1b – в первую очередь R1b-U106 и в меньшей степени R1b-P312. Таким образом «огузо-сакская» и «саксонская» R1b-компоненты расходятся уже на уровне древнейшего разветвления L23 → (Z2103 & M73) vs L51. R1b-Z2103 ассоциирован с ямной культурой (степи Причерноморья) и миграциями в Балканы, Кавказ (например, у армян и осетин). R1b-U106 связан с германо-скандинавскими миграциями (распространен в Северной Европе). Его формирование (~4 800 лет назад) произошло уже после разделения ветвей Z2103 и L51.
Их ближайший общий предок – R1b-L23 proto-ямной эпохи (~5400 лет до н.в.), то есть задолго до появления как тюркских, так и германских этносов.
Ниже перечислены опубликованные мужские образцы, которые стоят именно «на мостике» между базальным R1b-M269 и последующим расщеплением M269 → L23 → (Z2103 + L51). То есть они содержат либо: • M269+, L23– («pre-L23»), либо • M269+, L23+, но ещё не ушли ни в Z2103, ни в L51 – формальная запись L23*(xZ2103,xL51).
Для каждого скелета указано: код образца, памятник / культура, современная страна, калиброванная дата (2σ) и Y-SNP-статус.
EARLY ENEOLITHIC & ENEOLITHIC STEPPE (до протоямной)
• I0434 – Хвалынск II, мог. 101 (культура Хвалынск) Самарская обл., РФ │ 4550–4350 cal BC │ R1b-M269*(pre-L23)
• I5884 – Средний Потёмкинский кург. 4, мог. 1 (Eneolithic_Steppe) Оренбургская обл., РФ │ 4300–4000 cal BC │ R1b-M269*(pre-L23)
• I1171 – Progress-2, кург. 22/11 (кавказо-степной неолит) Ставропольский край, РФ │ 4300–4100 cal BC │ R1b-M269*(pre-L23)
STEPPE MAYKOP
• I6336 – Маринская 5, погр. 12 (Steppe Maykop) Кабардино-Балкария, РФ │ 3700–3520 cal BC │ R1b-L23*(xZ2103,xL51)
ЯМНАЯ (ранний горизонт)
• 1582.B (a.k.a. BZNK-1) – Базавлук I, кург. 16 (Yamnaya_Caucasus) Ростовская обл., РФ │ 3330–3010 cal BC │ R1b-L23*(xZ2103,xL51)
• I2104 – Гладковка, кург. 3 (Yamnaya_Lower Volga) Астраханская обл., РФ │ 3220–2880 cal BC │ R1b-L23*(xZ2103,xL51)
ПОЗДНИЕ «ЯМНО-КАТАКОМБНЫЕ» ПЕРЕХОДЫ
• I16168 – Майкопский р-н, кург. 5 (ранняя Катакомбная) Краснодарский край, РФ │ 2920–2570 cal BC │ R1b-L23*(xZ2103,xL51)
Сводка по датам 4550–4100 BC — первые M269*(pre-L23) в степи Волги и Предкавказья. 3700–3500 BC — первый надёжный L23* (Steppe Maykop). ≈3300 BC — L23* фиксируется уже в собственно ямных погребениях. <3000 BC — ветка раскалывается на Z2103 (базовая для «классической» Ямной) и L51 (будущие L51>P312/U106 в Европе).
Таким образом перечисленные семь образцов — это пока единственные опубликованные «общие предки» обеих основных линий L23 (Z2103 и L51) и, следовательно, предковое ядро для всей поздней R1b-ветви Ямной эпохи.
В то же время, до I тыс. н. э. ( со сдвигом, в первом тысячелетии нашей эры) все же найдены более восточные линии R1b (Z2103, M73, V1636, реже «базальные» L23*), тогда как «западно-европейская» ветвь L51 → (L151 → U106 | P312) на восток от Карпат археологически пока не прослеживается.
Ниже приведён перечень всех (на данный момент) единичных находок U106/P312-комплекса к востоку от Карпатского фронта
1) Причерноморье и Северный Кавказ: DA146 – Джейрах, Северная Осетия, культура Салтово-Маяк (алан/хуцар, VII–IX вв.)
R1b-L151>L52>L11(xU106,P312) – низкое покрытие, но явно уже в кластере L51.
• SMC001 – Самбек, Ростовская обл., то же горизонта Салтово-Маяк, VIII–IX вв.
R1b-U106>Z381>Z156 – интерпретируется как германо-аланский компонент среди кочевников.
• OLB003 – Ольвия, Николаевская обл., эллинистический некрополь, II в. до н. э.– I в. н. э.
R1b-P312>L2 – колонист из Средиземноморья.
2) Нижний Дунай / Балканы / Малая Азия
• VIM286 – Виминациум, Сербия, римский легионер, 2 в. н. э.
R1b-U106>Z381>P89.
• PLK201 – Велики-Преслав (Болгария), поздний First Bulgarian Empire, X в.
R1b-U106>Z381>Z156 – вероятно потомок западно-германских союзников/наёмников.
• LDC001 – Лаодикея-на-Ликосе (Турция), раннеримский некрополь, I в. н. э.
R1b-P312>DF27 – встроенный в римскую колониальную среду.
3). Иран и восточнее
• На сегодняшний день нет ни одного доновоевропейского образца с U106/P312.
В иранских, приаральских, тянь-шанских и алтайских выборках (Энеолит – Средневековье) фиксируются только Z2103, M73, R1a-Z93, Q1, C2 и др. «степные» линии.
4. Граничные/сомнительные случаи
• I2165 Yamnaya_Bulgaria (Гюргяни, ~3000 BC) – низкокавереджный L23*, первоначально предполагали, что это может быть «ранний L51», но точных downstream-SNP не прочитано.
• Пара низкокавереджных «R1b-L11?» в средневековых кладбищах Крыма и Причерноморья (объявлены в препринтах, но без подтверждённой фильтрации) – в публикациях 2023–24 гг. их либо переквалифицировали в L51, либо признали техническим шумом.
В качестве примера того, как линии скифов R1b заместили автохтонные славянские линии на Балтике, можно привести пример с Ольденбургской династией, потомков саксов, которая поселилась на исконной земле варягов и в их бывшей столице Стариграде, ныне Ольденбурге.
После обнаружения тел царя Николая II и его семьи в 1991 году живые двоюродные братья (Николай и Дмитрий) пожертвовали свои ДНК для тестирования Y-ДНК. Благодаря этому результату стала известна гаплогруппа Николая II (R1b), такая же, как и у датской королевской семьи. Семья Романовых вымерла после смерти императрицы Елизаветы 1-й в 1761 году. Новый царь Петр III происходил из дома Гольштейн-Готторпов. Это ветвь семьи Ольденбургов.
По предположению, учитывая, что все генеалогические связи одинаково биологические (без усыновления, без супружеской неверности), можно отнести эту же гаплогруппу ко всем членам семьи Ольденбургов мужского пола. Можно предположить, что все датские короли, начиная с Фридриха 1-го, родившегося в 1471 году и умершего в 1533 году, принадлежат к той же гаплогруппе, что и Николай II Российский, R1b.
Известные гаплогруппы правящих королевских семей:
Бельгия: R1b-U106 (Z305+)
Дания: до Фридриха IX: R1b, после Маргреты II: неизвестно
Испания: R1b-U106 (Z381+)
Люксембург : R1b-U106 (Z381+)
Норвегия: R1b
Соединенное Королевство: до Георга VI: R1b-U106 (Z305 +), после Елизаветы II: R1b
Показать полностью
2


Гунны-печенеги и половцы. Деполитизированная история
В настоящее время норманизм торжествует в исторической науке без каких-либо оговорок. Хотя давно доказано, что наиболее характерная мутация в Y-хромосоме у онемеченных славян, ставших скандинавами (шведами, датчанами, норвежцами) - это субклад R1a-Z284. Он принципиально отличается от субклада R1a-Z280 у восточных славян и R1a-M458 западных славян, к которым принадлежали варяги.
Несмотря на научно доказанную несостоятельность норманизма, в 2012 г., когда праздновалось 1150-летие зарождения российской государственности, археолог Сергей Щавелёв публично поделился аксиомой норманизма: небольшая группа викингов, несомненно обладавшая более сложной культурой и менталитетом лидеров, во главе с Рюриком основала русское государство. Как он пишет в речит-лекции, посвященной зарождению российской государственности, финальный этап славянского этногенеза археологи и лингвисты связывают с серединой I тысячелетия новой эры. Это время, когда на территории Европы проживали многочисленные племена, являющиеся общими предками будущих славян. Восточные славяне имели соседями на севере финно-угров и балтов. На юге их соседями были потомки будто бы ираноязычных (на самом деле тюркоязычных) сарматов — аланы (одни из предков немцев), а также болгары. Кроме того, восточные славяне соседствовали с новыми кочевыми народами, такими как авары (обры, предки бавар и лангобардов), мадьяры (угры, предки венгров), печенеги (гунны из Семиречья к северу от Аральского моря, также предки немцев), аланы (ясы, предки немцев и венгров) и половцы (кипчаки из степей между монгольским Алтаем и восточным Тянь-Шанем, также предки немцев).
Господство норманизма в России привело к тому, что бывший премьер-министр Швеции Карл Бильдт, писал в «Твиттере»: «Это был античный греческий город Херсонес, где викинг, князь киевский Вальдемар был крещён. Сомнительные основания для Москвы претендовать на Крым».
Хотя Херсонес, ранее называвшийся Корсунью, Хорсунью в честь культа Хорса (Христа, писалось как ХРС в во времена патриарха Фотия), был основан как русский город, филиал столицы Всея Руси Ладоги (33 вторичной Арконы, Куявы, Куябы, Киявы, Кия, Стрелы Яра, Киева) в 861 г. Херсонес был главным портом Черного моря наряду с филиалом Арконы Белгород Днестровским (по-турецки Аккерман - «Белый камень»). В связи с княжескими междоусобицами на Руси, Херсонес в 109 г. до н.э. (1155 г. н.э. с учетом сдвига) был завоеван царем Понта Митридатом VI Евпатором при участии готов, которые прежде, в конце III в. (IXв.), взяли под контроль Понтийское царство (281 г. до н.э. - 62 г. н.э., или 983 - 1326 гг.) и встроились в его элиту, сохраняя традиции военной демократии. Между тем, вскоре в Крым пришли новые переселенцы - тюрки.
Тюркский каганат, XI - XII вв.
Огромное влияние на всю мировую историографию оказали тюрки, заселившие к тому времени вместе с аланами и готами всю Европу. Их государство так и называлось - Тюркский каганат, который сформировался в VI (с учетом сдвига в XI) веке на территории Алтая в составе Всея Руси.
Первоначально это был союз племён, принявших название «тюрк» или «тюркют». Территориально каганат располагался на обширных землях от Волги и Аральского моря до Северного Китая. В состав каоаната входили:
Территория современного Казахстана
Согд и Бухар
Северный Кавказ (Алания
Подонье (Русколань)
Каганат просуществовал до 603 (XII века с учетом сдвига) года, когда распался на Западно-тюркский и Восточно-тюркский каганаты из-за междоусобных войн и противостояния с Византией, Ираном и Китаем.
В 560-х (VIII в.) годах тюрки вышли к Волге, Северному Кавказу, Дону и побережью Азовского моря, где эти территории принадлежали савирам (сарматам), которые также назывались русами (роксаланами Руси Лоновой, Русколани) и аланами-ясами (выходцами с Кавказа, предками осетин).
Аланы были одним из основных народов, попавших в зависимость от Тюркского каганата в волго-донских степях. Это подтверждается словами тюркского правителя, записанными византийскими послами в 570-х (XI в.) годах:
"Посмотрите, несчастные, на аланские народы... которые были одушевлены безмерной бодростью, полагались на свои силы и осмелились противустать непобедимому народу тюркскому, но они были обмануты в надеждах. Зато они в подданстве у нас, стали нашими рабами".
Отношения между тюрками и аланами изначально были враждебными, но не слишком ожесточенными. В отличие от событий конца IV (X) века, в VI (XII) веке в Приазовье не было масштабных разрушений и жертв, что связано с тем, яс Один вместе с союзными племенами ясов покинул Асгард - Азов в поисках лучшей доли.
В 370-380-х (на самом деле, в X веке) годах гунны-печенеги прошли мимо Крыма и обрушились на «готское государство» Германариха, начав теснить союзы алан и готов. В течение второй половины V и начала VI века (XI-XII вв.) над Боспором распространялся «протекторат» гуннского племени утигуров, вернувшихся из Европы после распада Гуннского союза. Надписи с именами царей династии Тибериев-Юлиев датируются вплоть до конца V (XI) века.
В 520—530-х годах (XI в.) Византия устанавливает над Боспором прямую власть. Будто бы античный период, а на самом деле, русский период его истории плавно переходит в византийский без разрывов в эволюции материальной культуры. В 576 (XI в.) году территорию от современной Грузии и до Крыма завоёвывает после войны с Византией Тюркский каганат - союз гуннов-огузов.
В ходе Третьей Митридатовой войны, начавшейся в 74 году до н. э. (1190 г. по альтернативной хронологии), римский полководец Гней Помпей Великий одержал ряд побед над Митридатом VI Евпатором, что вынудило последнего бежать в Крым. В 63 году до н. э. (1201 г. по альтернативной хронологии) Митридат, опасаясь пленения римлянами, покончил с собой в Пантикапее (современная Керчь).
После смерти Митридата его сын Фарнак II предпринял попытку восстановить утраченные владения отца в Малой Азии. Оставив Асандра наместником на Боспоре, Фарнак направился в Азию. Однако к тому времени Гай Юлий Цезарь уже одержал победу над Помпеем и двинулся навстречу Фарнаку. В битве при Зеле (современный турецкий город Зиле) в 47 году до н. э. (1217 г. по альтернативной хронологии) Цезарь разгромил армию Фарнака. В честь этой победы Цезарь произнёс знаменитую фразу: «Veni, vidi, vici» (Пришёл, увидел, победил). В результате Понтийское царство (983 - 1326 гг.), также известное как Трапезундская империя (1204–1461 гг.), было завоёвано и присоединено к Римской империи. В 25 году до н. э. (1239 г. по альтернативной хронологии) Октавиан Август предоставил Херсонесу статус «свободного города», освободив его от власти боспорского царя и подчинив непосредственно Риму. Однако римская экспансия на Чёрном море вскоре столкнулась с противодействием кипчаков-скифов Яра (основное население Золотой Орды), что привело к разрушению римской Корсуни ханом Ногаем в 1278 г.
В 60-е годы I века (1320–1330 годы по альтернативной хронологии) нашей эры римляне провели крупную военную кампанию в Таврике для противодействия скифам. Легат провинции Мезия Плавтий Сильван успешно возглавил эту экспедицию по поручению императора Нерона и одержал победу над скифами. В результате Херсонес стал важным стратегическим форпостом римских вооружённых сил в Северном Причерноморье и фактически вошёл в состав провинции Нижняя Мезия.
Как мы видим на примере Крыма, то есть "Кромки" Скифии Яра и Скифии Мары, причиной локального национализма в различных странах Европы, предающего забвению русскую историю и русскую принадлежность освоенных территорий, в том числе и Швеции, священной для норманистов, является потребность застолбить пришлыми племенами славянские территорий Европы в ходе Великого переселения народов. В III веке (с учетом хронологического сдвига примерно в 600 лет, в IX веке) готы с характерной гаплогруппой I1, продвигаясь с боями, пересекли территории от Готского полуострова через Поморье, Прибалтику, Польшу и Малороссию, столицей которой была крепость Киев (Стрела Яра) на Днепре, до Северного Причерноморья и Италии. Попутно готы вступали в военные союзы с аланами, лангобрдами и пр., сохранив, тем не менее, русское самоназвание венды-вандалы. Как ни странно, согласно картам римского историка Тацита (1332 - 1396 гг.), ругии (руяне, арконцы), варины (варяги) и готоны (готы) еще существовали как отдельные этносы вплоть до XIV века. А если мы обратимся к хронистам франков, которые начали править Европой лишь с XIV в., то эти этносы существовали вплоть до XVII в., что переводит нас к относительно недавним событиям попытки России воссоединения с Пруссией, которую саботировал чистокровный сакс Петр III.
В 375 (942, сдвиг 567 лет) г. гунны Гунгарии разгромили Готское царство Германариха (265-375 или 832-942 гг.). В I книге "Войны с готами" Прокопий Кесарийский описывает этнический состав войск византийской армии под началом Мартина и Валериана, выступивших против остготов из италийского Королевства остготов (просуществовало в 489-555, или 1056-1122 гг. со столицей в Равенне) в 538-539 гг. (или 1105-1106 гг.) :
"гунны, славины и анты, которые живут по ту сторону Дуная, недалеко от его берегов".
Первый цезарь Руси Яровой Римской (Римской республики), Феодосий I Великий (347 - 395 гг., или 955 - 1003 гг.) успешно отразил их первую волну, подгоняемую гуннами, в Готской войне (377-382, или 985-990 гг.). Вместе с тем столь тесное общение с оседлыми народами не могло не отразиться и на самих печенегах. С изменением социально-экономической структуры их кочевого общества, единый когда-то союз печенежских племен с 20-х гг. XI в. начинает распадаться на отдельные орды. Очевидно, не последнюю роль в этом процессе сыграло появление в Восточной Европе новой волны тюркоязычных кочевников – половцев-кирчаков, потомков огузов (40-е гг. XI в.).

Аттила на троне. Chronicon Pictum, факсимильное издание, хранящееся в библиотеке Университета Мэриленда.
В битве на Каталаунских полях Галлии, состоявшейся под Труа (современная Шампань во Франции) в 451 (1018) г., участвовали объединенные войска Римской Республики и Тулузского королевства вестготов против союза гуннов (печенегов, кунгаров) и немецких племен под началом Аттилы, который жил в 395-453 гг., или 962-1020 гг. и стал единоличным властителем гуннов в в 434 г. - 1001 г. (сдвиг 567 лет для гуннов, печенегов). Если Аттила жил в 962-1020 гг., то его двойником является Иштван I (997-1038) - первый коронованный король Венгрии (с 1001 года).

Останки Аттилы находятся в приделе Базилики святого Иштвана. Речь идёт о мумифицированной правой кисти короля Иштвана - Священной деснице (Szent jobb).
Миграции с участием вышеперечисленных эрбинов (представителей гаплогруппы R1b) на исконно русские территории подтверждаются тем, что, например, в Саксонии-Анхальт, то есть в центре Германии, жил в 1113-1021 гг. до н. э. русскоговорящий серб-лужичанин с характерным субкладом восточных славян R1a-Z280. А Сербия говорит на диалекте русского языка.
Согласно историческим данным, первоначальные территории обитания печенегов (гуннов) были связаны с бассейном Сырдарьи и районом от современного Ташкента в сторону Хорезма, то есть неподалеку от места зарождения гаплогруппы R1b. До конца IV (X) века гунны-печенеги проживали в районе между Давани (Фергана) и владениями Яньцай (Аланья). С конца IV (X) по начало VII (XIII) века, находясь под властью эфталитов (белых гуннов), они соседствовали с огузами и вели постоянные войны за пастбища вокруг иссыхающего Аральского моря. Именно здесь, в бывшем Семиречье, зарождалось ядро немецкого этноса.
Существенный вклад в этногенез немцев, особенно саксов, внесли и обитавшие здесь саки. Саки считаются потомками носителей древностей срубной и алакульского варианта андроновской культур. Иранские источники называют саков «турами с быстрыми конями». Древнегреческие авторы называли саков «азиатскими скифами», то есть скифами Мары. После завоевания Персидской империи Александром Македонским (857-889 гг. жизни, сдвиг 1213 лет) в 334-331 гг. до н.э. (879-882 гг. н.э.), одна из групп сакских племён вторглась в пределы сатрапии Селевкидов Парфиены (III век до н. э.) и сыграла значительную роль в образовании Парфянского царства в 247 г. до н.э. (966 г. н.э.): вождь племени парнов Аршак стал основателем парфянской династии Аршакидов. Первой столицей Парфии стал Гекатомпил в районе современного Шах-Руд (также известного как Шахри-Кумис) около города Дамган в Иране.
Часть племён саков вторглась в Дрангиану и заняла её, дав ей название Сакастан (Страна саков, у Страбона лат. Sacasene - современный Систан на территориях Ирана и Афганистана).
Страбон достаточно подробно описывает набеги саков, сообщая, среди прочего, что они добирались даже до Каппадокии. Переселение саков было частью перемещения кочевых народов, запущенного вытеснением юэчжей племенем хунну около 175 г. до н. э. (1038 г. н.э.). В итоге они вместе с потомками первой волны переселения народов данами добрались до Саксонии и, к величайшей радости Рим, разграбили столицу Руси Яра - Аркону.
Показать полностью
3