Вы спрашивали о палеонтологии.
У Лиги Палеонтологии день рождения, и вчера мы попросили вас задать вопросы о том, что бы вы хотели узнать о палеонтологии. Орфография вопросов сохранена)
Вопрос:
Палеонтология на других планетах – фантастический миф или возможная реальность?
Отвечает @MartinDont:
Поиск жизни на других планетах, следов её прибывания и изучения её возможной эволюции (что по сути частично является космопалеонтологией) не просто возможная реальность, а сегодняшний проект NASA. Цель программы NASA по экзобиологии и эволюционной биологии заключается в том, чтобы понять, как произошла жизнь, какова была её дальнейшая эволюция, как она распространялась во Вселенной и какое у нее будущее. Исследования сосредоточены на происхождении и ранней эволюции жизни, на её потенциальной адаптации к различным средам, и изучении того, какие могут быть последствия для жизни в условиях, отличающихся от земных. Эти исследования проводятся в составе текущего проекта NASA по изучению нашей звёздной системы и определении её биосигналов на Земле. Никакой фантастики
https://hi-news.ru/science/kak-zhizn-stala-slozhnoj-i-mozhet...

Вопрос:
Если бы один миллион лет назад на Земле вымерла какая-нибудь разумная цивилизация, сравнимая по развитости с нынешней, то какие свидетельства их разумности могли бы дойти до нас?
Отвечает MartinDont:
Развитая цивилизация подразумевает хорошо организованный социум с налаженным производством предметов потребления. Хоть миллион лет назад, хоть десять, но следы подобных цивилизаций остались бы в земле в виде культурного слоя в огромных количествах. Огромные промышленные комплексы из стали (а любая цивилизация так или иначе должна использовать столь практичный и легкодобываемый материал как сталь) и прочих металлических сплавов обязательно оставили бы свои следы в недрах Земли. Инструменты, отходы, окаменелые остатки сельхоз животных и сами останки людей не исчезают бесследно даже за миллионы лет. В 2016 году были найдены примитивные каменные орудия возрастом более миллиона лет, что ж уж говорить о промышленных комплексах. http://paleonews.ru/new/844-homovs

Вопрос:
Какова вероятность найти образец, который не будет вписываться в существующее дерево эволюции?
Отвечает MartinDont:
Ежедневно находятся новые виды, роды и даже семейства, которых ещё нет на эволюционном древе. Постоянно проходят таксономические исследовании, перемешивающие родословную организмов. В 2018 году стартовал проект по изучению филогении птиц, что сулит большими сюрпризами. Открываются новые замкнутые экосистемы, где организмы обособленно от остального мира эволюционировали миллионы лет, как например чёрные курильщики. Чёрные курильщики – это такие гидротермальные источники, выбрасывающие воду температурой 350-400 градусов в океан под большим давлением. Сама вода перенасыщена тяжёлыми металлами. На удивление биологов, чёрные курильщики оказались настоящими оазисами жизни. Организмы вокруг курильщиков не только приспособлены к экстремальным условиям, но и живут без света вовсе, а питаются серой.
А если о говорить о невероятных организмах, то шанс найти что-то подобное мал. Мы уже имеем хорошее представление о том, как протекает эволюция. Сильно удивить нас могут замкнутые экосистемы и их обитатели.
Также мы можем найти новые группы животных, о которых мало что знаем, и которые буду сильно выделяться на фоне уже изученных организмов (таких, как вендобионты https://ru.wikipedia.org/wiki/Проартикуляты). Их неясное положение в эволюционном древе — временно. Всё встанет на свои места, как только мы получим достаточное количество ископаемого материала.
Вопрос:
Является ли любительская палеонтология легальной? Планирую поехать в Ундоры, посоветуйте пожалуйста, как найти что- нибудь интересное, и какие инструменты стоит взять?
Отвечает: PaleoHunters.ru
Понятие любительской палеонтологии очень неточное. Проводить раскопки без лицензии категорически запрещено. Карается подобное огромными штрафами в сотни тысяч рублей и тюремным заключением. С другой стороны, поверхностный сбор разрешён. Можете спокойно собирать аммонитов и трилобитов в карьерах, что лежат на самой поверхности. И вновь же, с другой стороны, поверхностный сбор без лицензии запрещён в заповедных зонах, коих очень много в Ульяновской области. Уголовных и административных статей для наказания чёрных копателей очень много.
https://ru.wikipedia.org/wiki/%D0%9C%D0%B5%D0%BB%D0%BE%D0%B2...
Вопрос:
Сколько мог жить динозавр?
Отвечает MartinDont:
Сложно судить о сроках жизни животных, чей метаболизм для нас неизвестен. Но у нас есть годичные кольца на костях. Разные динозавры взрослели по разному. Например, тираннозавр в 20 лет становился взрослой особью, а знаменитый тираннозавр Сью в свои 28 лет была уже старушкой. Гадрозавры переходили во взрослую стадию в 15 лет. Гигантским зауроподам с их предположительно вялым метаболизмом дают десятки лет жизни, а небольшим и активным дромеозавридам всего 10-15 лет.

Вопрос:
Как появилась жизнь на земле?
Отвечает DragonSpace:
Для начала стоит ответить на вопрос: когда появилась жизнь?
В сентябре 2015 года национальная академия наук США выложила результаты исследования изотопов углерода, в которых, по мнению учёных, были обнаружены следы биологической активности. Этому углероду более 4 миллиардов лет. Наша планета недолго оставалась одинокой и примерно через 500 млн лет после зарождения самой Земли, зародилась и жизнь на ней.
Теорий появления жизни на Земле великое множество, но рассмотрим наиболее вероятную.
У нас три основных критерия для зарождения жизни:
- наличие определённых металлов в концентрациях, превышающих фоновые (цинк, кобальт, марганец),
- наличие фосфора, без которого цепочки ДНК и РНК не возникли бы
- солнечный ультрафиолет,
- наличие воды, но в меру.
Такому набору условий чётко отвечает только горячие источники вблизи вулканов, а именно так называемые грязевые котлы, или фесселиты.
Правда, современные грязевые котлы содержат много серной кислоты, которая получается при реакции сероводорода с кислородом воздуха, и практически необитаемы. Но в древние эпохи, когда кислорода в атмосфере еще не было, грязевые котлы должны были иметь нейтральную среду и быть пригодными для жизни.
А при ближайшем рассмотрении этих котлов оказалось, что они предоставляют почти всё необходимое для зарождения жизни:
• среду, обогащенную калием, фосфором и необходимыми микроэлементами;
• местообитание со встроенным источником тепла, с практически постоянными условиями независимо от капризов погоды;
• пористые минеральные осадки, работающие в качестве катализаторов и предоставляющие огромное количество раздельных микроотсеков для обитания доклеточных форм жизни;
• испаряющиеся лужи, в которых могут накапливаться органические вещества и благодаря высокой концентрации солей и формамида может идти образование цепочек РНК и белков,
Никакие другие местообитания не обладают сразу всеми этими достоинствами. Например, в «курильщиках» нет обогащения калием и фосфором, нет ультрафиолета и нет накопления веществ в испаряющихся лужах. Так что грязевые котлы наземных геотермальных полей на сегодня представляются самым вероятным местом появления жизни.
Ответ преимущественно взят у @2diesel, который отлично законспектировал книгу М. Никитина Происхождение жизни. От туманности до клетки".

Вопрос:
Как появились млекопитающие?
Отвечает DragonSpace:
Млекопитающие произошли от Синапсид, древних четвероногих, процесс их появления называется маммализация териодонтов(один из множества эволюционных ветвей синапсид). Появились млекопитающие после пермь-триасового великого вымирания примерно 225 млн лет назад, напомню, что динозавры появились примерно 230 млн лет назад. Первые млекопитающие всё ещё отдаленно напоминали рептилий, яйцекладущие, без развитого мозга, но уже с активной терморегуляцией, и 4хкамерным сердцем.
Молочные железы возникли из потовых желез, и изначальная функция была скорее в выпаивании детенышей, чем в выкармливании.

Вопрос:
Рыбы - монофилетический таксон или нет?
Отвечает DragonSpace:
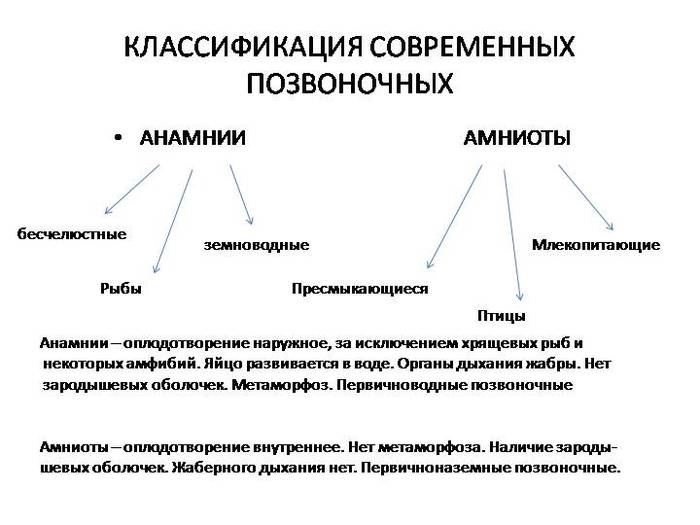
Чтобы зваться монофилетической, группа должна включать предка и его потомков.
Таксон "Рыбы" не является монофилетическим, так как не включает амфибий и амниот.
Вопрос:
Был ли у вас когда нибудь страх, что при исследовании какого-нибудь нового, неисследованного вида условного трилобита тот окажется в состоянии сохранять энергию в режиме "гибернации", а после пробуждения накинется на вас а-ля лицехват?
Отвечает DragonSpace:
Жизнь строится по усложнению, от примитивных существ, к более прогрессивным. Чем раньше жил какой-либо вид, тем он вероятно более примитивен(идея тем точнее, чем больше между двумя сравниваемыми видами прошло времени).
И так как до сих пор ни у кого на планете не обнаружены такие необычные способности, никто не ожидает найти трилобита в гибернации.
Вопрос:
Можно ли сказать, что мы состоим из миллиарда других живых существ (людей, живших ранее, динозавров, насекомых и тд)?
Отвечает DragonSpace:
Можно сказать, что мы состоим из миллиарда тех же атомов, которые раньше были частью других живых организмов. Как говорил Муфаса: Все мы связаны в великом круге жизни.
Вопрос:
Скажите пожалуйста, что есть нефть? Всё таки динозавры или планктон?
Отвечает DragonSpace:
Доминирующая биогенная теория гласит, что нефть и природный газ образовались из остатков растительных и животных организмов в ходе многоступенчатого, длящегося миллионы лет процесса. Сырьём для образования нефти на 99.99% служили остатки зоопланктона и водорослей. Подробнее:
https://pikabu.ru/story/proiskhozhdenie_nefti_trupyi_dinozav...

Вопрос:
Почему до ледникового периода зверье было таких огромных размеров? И после ледникового, так и не появилось больше крупных животных, кроме китов, даже в местах с благоприятным климатом, где нет зимы и много растительности.
Отвечает DragonSpace:
На нашей планете то и дело происходили различные катаклизмы, которые вели к освобождению и перестройке экологических ниш. Каждое животное становилось огромным, потому что осваивая новую нишу, приходилось жестко адаптироваться к меняющимся условиям, так, Мезозой славился гигантизмом динозавров, а Кайнозой – эра правления млекопитающих. Гигантизм сам по себе вполне естественный эволюционный путь доминирующей группы животных – млекопитающих в данном случае. Конкретно механизм гигантизма развивается через соревнование по выживанию хищника и его жертвы – одни растут, чтобы лучше защищаться, вторые, чтобы догнать первых и проще убивать, в конце концов такая гонка приводит самих животных в тупик, тк те же самые экологические ниши продолжают меняться и распадаться, а гигантам приспосабливаться к новым условиям очень сложно.
К слову, последний ледниковый период не закончился, закончилась ледниковая эпоха ледникового периода, сейчас же межледниковье ледникового периода.
Если брать гигантов, живших недавно, то человек, своим расселением по планете уничтожил множество видов, большинство из которых и так были на грани вымирания, но с тех пор прошло слишком мало времени, для того, чтобы ниши заполнились заново.
А если брать весь кайнозой в целом, то исчезновению гигантов способствовала два ключевых фактора: резкое изменение условий окружающей среды – катаклизм, и стратегия родственной стайности оказалась намного эффективнее гигантизма, и успешно его вытеснила.
Поэтому последний ледниковый период/эпоха не заслуживают такого отношения к себе, ведь и сейчас на земле полно гигантов – два вида слонов, носороги, жирафы, киты и главный рекордсмен - синий кит, а самое большое сухопутное животное Индрикотерий существовал задолго до последнего похолодания.

Вопрос:
Черепахи и крокодилы выжили после падения метеорита (их ближайшие предки) благодаря панцирям и чешуе?
Отвечает @p4hshok:
Черепахам и крокодилам нужно гораздо меньше пищи чем динозаврам.
За один раз крокодилы могут съесть до 23 % от массы своего тела. До 60 % съеденной пищи у крокодилов может переходить в жир, запасаемый в специальных полостях между мышцами, что позволяет им переживать длительные периоды голодовки. Будучи холоднокровными животными, крокодилы требуют примерно в пять раз меньше пищи, чем теплокровные хищники такого же размер. Крокодилы хорошо адаптированы для длительного голодания. Без еды взрослые крокодилы в случае крайней необходимости могут обходиться примерно до одного года. Даже только что вылупившиеся детеныши за счет понижения уровня обмена веществ способны прожить без пищи около 58 дней, потеряв при этом 23 % от своей массы.
Вопрос:
Ближайший общий предок всех членистоногих - кто он? От кого произошли насекомые?
Отвечает p4hshok:
Главная проблема последнего общего предка всех членистоногих в том что его может просто не быть, так как на границе венда и кембрия артроподизация https://bio.wikireading.ru/3905 (накопление признаков членистоногих) происходила параллельно и достоверно определить предка членистоногих невозможно потому что их может быть несколько.
Палеонтологическая летопись так же не даёт ответа так как в кембрии у нас уже есть широчайшее разнообразие членистоногих, а в венде только вендобионты родственное отношение которых к многоклеточным животным крайне спорно.
Сприггина например вполне напоминает трилобита, но симметрия скользящего отражения (https://ru.wikipedia.org/wiki/Скользящая_симметрия) ставит учёных в тупик и заставляет усомниться в родственности вендобионтов к двустороннесимметричным животным.
Есть попытки реконструировать его внешний облик: Последний общий предок членистоногих реконструируется как сегментированный организм, каждый сегмент которого покрыт собственным склеритом и несёт пару конечностей. Вопрос о типе этих конечностей остаётся открытым. Это прачленистоногое имело на брюшной стороне рот, а на передней части спинной — глаза. Антенны были расположены перед ртом. Питалось оно, вероятно, пропуская через себя донные осадки.
Вероятнее всего общий предок вех членистоногих, если вообще существовал, должен быть из позднего докембрия небольшого размера да ещё и без полноценного твёрдого покрова.
Новейшие морфологические сравнения и филогенетические реконструкции на основе геномных последовательностей указывают, что насекомые являются потомками ракообразных, а не сестринским таксоном. Это заключение хорошо согласуется с палеонтологическими данными. Однако морфологические и молекулярные данные не согласуются при определении ближайших родственников насекомых среди ракообразных: морфологические данные указывают на связь насекомых с высшими ракообразными, а молекулярные — с жаброногими.
В соответствии с последней гипотезой эволюционная ветвь насекомых отделилась от ракообразных в позднем Силуре — раннем Девоне. Эта оценка согласуется и с палеонтологическими данными, и с оценкой на основе молекулярных часов.
Так же лекции про эволюцию членистоногих:
https://www.youtube.com/watch?v=csLewT3aMb8
https://www.youtube.com/watch?v=-OC46Vjg0jU

Вопрос:
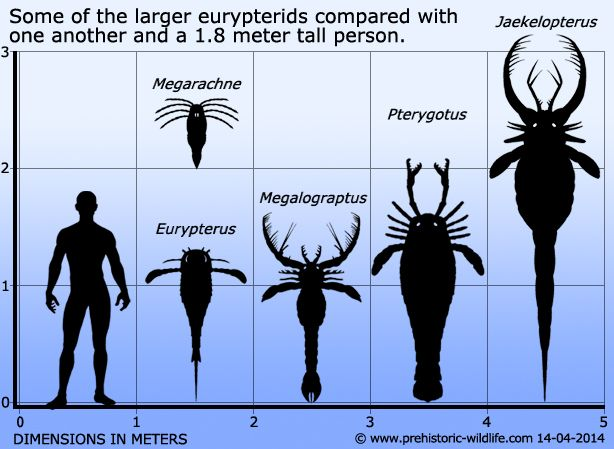
Почему вымерли ракоскорпионы и трилобиты?
Отвечает p4hshok:
Потому что были вытеснены более прогрессивными видами, а также подверглись давлению нескольких массовых вымираний. Рыбы обзавелись панцирями и челюстями стали конкурировать с ракоскорпионами и даже охотиться на них постепенно вытесняя их в мелкий размерный класс. К каменноугольному периоду остались лишь пресноводные виды. Окончательно вымерли во время великого пермского вымирания.
С трилобитами та же история архаичные и некогда многочисленные они так же не выдержали давления новых прогрессивных видов и массовых вымираний.
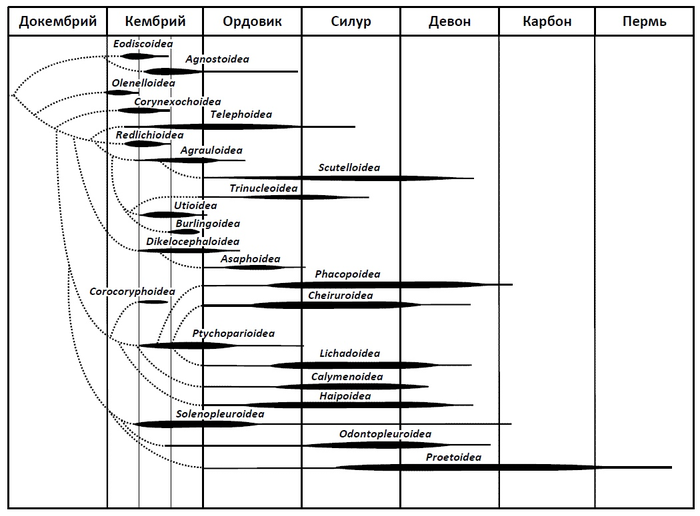
До перми доживает только один подкласс мелкоразмерных Проетида https://en.wikipedia.org/wiki/Proetida Обрывается славная история трилобитов так же пермским вымиранием.
Разнообразие трилобитов в геохронологической шкале.
Вопрос:
Я читал, что в процессе эволюции природа в короткий срок создаёт множество видов, потом в течение некоторого времени большинство их вымирает, остаются самые приспособленные. подсчитывали ли учёные КПД эволюции с точки зрения расхода ресурсов?
Отвечает p4hshok:
Нет никому это не нужно, вещества находятся в круговороте и интересно это может быть только математикам для построение соответствующей модели, чисто для тренировки мозгов.
Вопрос:
Есть ли виды которые пережили ледняковый период и как им это удалось?
Отвечает @AntonPerm
Тут нужно уточнить, какой именно ледниковый период имеется ввиду, так как их было насколько, но скорее всего, как я понимаю, имеется тот, который показан в мультике. На самом деле, правильно этот этап оледенения называется ледниковая эпоха четвертичного ледникового периода. Эта эра закончилась на столь давно, примерно 9000 лет назад, поэтому большинство животных, которых мы видим сейчас, можно сказать, пережили это похолодание, проще назвать тех, кто не пережил его, например, шерстистые носороги. Даже последние мамонты вымерли 3500 лет назад, а значит и они пережили эту эпоху. Ну и, конечно, человек, как вид, превосходно пережил это великое оледенение.

Вопрос:
Реально ли собрать небольшую экспедицию в ту же Азию, порыть-поискать кости дино и, в случае находок, перевезти через границы и оставить их себе?
Отвечает AntonPerm:
Азия большая, и очень сильно насыщена окаменелыми остатками динозавров, расскажу на примере 2 стран - Монголии и Казахстана. В Монголии очень строгие законы касаемо раскопок, туда не пускают обычных туристов, а если приглашают палеонтологов из-за рубежа. то вывозить находки строжайше запрещено, можно даже принудительно остаться в Монголии на несколько лет дольше запланированного. В Казахстане-же, Вас с удовольствием свозят на меловой карьер, где можно поискать разнообразные окаменелости. Как дело происходит в Китае, если честно не скажу, но мне кажется, что примерно как в Монголии. а

Вопрос:
Что познавательное и интересное можно на эту тему посмотреть детям-дошкольникам?
Отвечает AntonPerm
Для дошкольников отлично подойдёт мультфильм "Поезд динозавров", там и описаны приключения героев и даны комментарии палеонтолога. Дошкольникам постарше можно включать фильмы BBC наподобии "Прогулок с динозаврами"

Вопрос:
как вообще найти какую то окоменлость?
Отвечает AntonPerm
Для этого нужно изучить геологическую карту местности, на которой Вы собираетесь проводить раскопки, тщательно подготовить специальные инструменты и изучить ту фауну и флору, типичную для данной местности того периода, ну, или просто внимательнее смотреть вокруг, а особенно под ноги. Зачастую окаменелые остатки можно найти даже на газоне. Ну и конечно, рекомендую посетить карьеры, как правило, там можно найти много интересного.

Вопрос:
Жена выедает мне мозг, чтобы я завёл в вк паблик с названием "Говно мамонта" и выкладывал там палеонтологические байки и занимательные факты. Стоит ли мне это сделать?
Отвечает AntonPerm
Почему бы и нет) Палеонтология распространена по всему миру и повсюду происходит очень много любопытного и весёлого, поэтому почитать об этом будет весьма интересно. А начать можно с выкладывания историй в нашу "Лигу палеонтологии", посмотреть, так сказать, реакцию публики.

Вопрос:
объясните, почему раньше всё так классно окаменевало, а теперь тупо гниёт без следа?
Отвечает AntonPerm
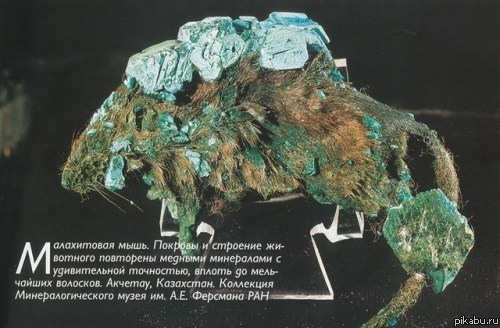
Нет никакой разницы между сохранностью организмов тогда и сейчас. Очень многое что сгнило много миллионов лет назад мы уже никогда не найдём. Чтоб до нас дошли окаменелые остатки животных и растений должно сложиться множество факторов. Например если динозавр умер в лесу, то, скорее всего, природа переработает его тело чуть менее чем полностью, но если тот самый динозавр умрёт в пустыне или в болоте, то мы сможем увидеть то, что от него останется. Если перенестись на 60 миллионов лет вперёд, то мы обнаружим остатки современных животных примерно в том виде, в котором мы находим динозавров.
В пример можно привести знаменитую малахитовую мышь, попав в подходящие условия органика довольно быстро заменилась минералами, теперь можно назвать эту мышь окаменелостью.
Вопрос:
Расскажите, пожалуйста, как вот столько рыб сразу отпечаталось?
Как вообще отпечатываются существа в породах?
Да, я тупой несмышленыш
Отвечает AntonPerm
Как уже говорил выше, для того чтоб до нас дошли окаменелые остатки, должно сложится много факторов. В данном случае, скорее всего, рыбы во время разлива заплыли в заводь, которая при отливе стала отрезана от реки, там быстро кончился кислород, и весь косяк упокоился на дне, где и был занесена илом.
Примерно так и проходит фоссилизация(окаменение), за много лет органика замещается на неорганику(псевдоморфоз), и те кости, которые мы видим, по сути являются камнем по химическому составу.

Ответили почти на все вопросы, если что то осталось непонятным, добро пожаловать в комментарии)