Талос - род североамериканских троодонтид, живший 70 млн лет назад
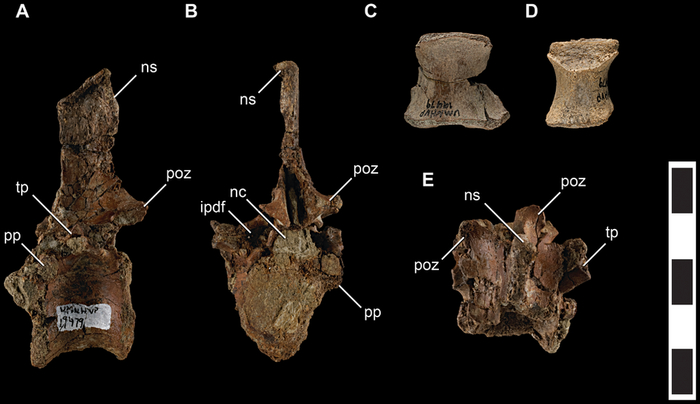
Талос известен только по образцу UMNH VP 19479, частичному посткраниальному скелету субадультной особи, включающему фрагменты передних конечностей, таз, фрагменты позвонков, шевроны и левую локтевую кость. Он был обнаружен в 2008 году М. Дж. Кнеллом во время проекта "Котловина Кайпаровиц", инициированного Университетом штата Юта в 2000 году, в формации Кайпаровиц в Национальном памятнике Гранд Лестница-Эскаланте.
Восстановленная нога
Впервые талос был назван Линдси Э. Занно, Дэвидом Дж. Варриккио, Патриком М. О'Коннором, Аланом Л. Титусом и Майклом Дж. Кнеллом в 2011 году, типовым видом является Talos sampsoni. Родовое название образовано от имени героя древнегреческой мифологии, Талоса, гигантского бронзового витязя, с включением английского слова "talon", "коготь". Видовое название дано в честь палеонтолога доктора Скотта Сэмпсон за изучение и сбор окаменелостей в рамках проекта "Котловина Кайпаровиц".
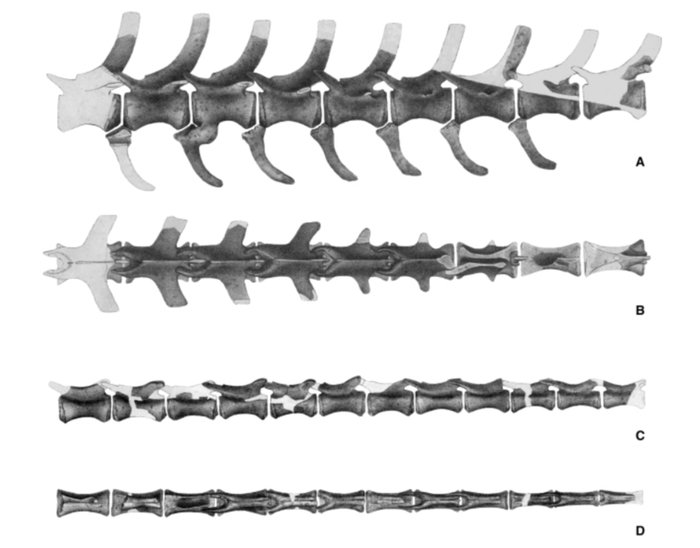
Позвонки
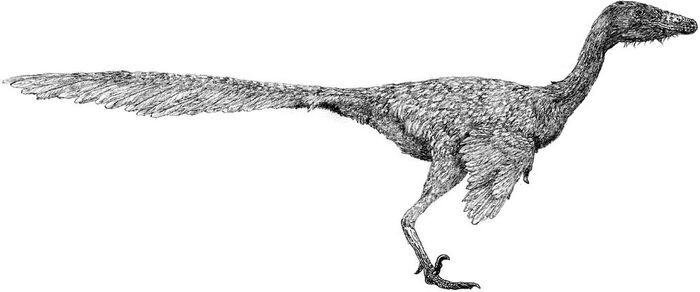
В длину талос достигал 2 метров, а его вес оценивается в 38 килограмм. Черты животного более элегантны, чем у Troodon, но чёткого представления о облике Talos sampsoni нет в связи с фрагментарностью находки. Вероятно, внешне он был похож на других троодонтид, обладая короткими передними конечностями и длинными ногами. Определенно точно, это был теплокровный двуногий динозавр, покрытый перьями. Особенностью известного образца является частично поврежденный серповидный коготь левой ноги, несущий следы заживления. Авторы описания предполагают, что палец ноги мог быть поврежден в результате какой-либо физической травмы или инфекции. Согласно популярной теории, он был поврежден в результате охоты, хотя это остается спекулятивной идеей.
Реконструкция
В 2011 году талос был определен в семейство троодонтид. Кладистический анализ показал, что он входил в кладу производных троодонтид вместе с Byronosaurus, Saurornithoides, Zanabazar и Troodon.
Talos sampsoni вместе с Troodon — первые представители троодонтид из верхнего мела Северной Америки. В те времена там был тёплый и влажный климат. По описанию нового динозавра учёные предположили, что, вероятно, в те времена там обитало много неописанных видов, близких к троодону. Эта гипотеза связана с очень широким пробелом (20 миллионов лет), к которому можно отнести множество остатков, представляющие собой плохой и фрагментарный материал.
Единственный известный образец талоса был обнаружен в формации Кайпаровиц в южной части штата Юта. Радиоизотопное датирование показывает, что формация Кайпаровиц была отложена между 76,1 и 74,0 млн лет назад, во время кампанского яруса мелового периода. Тогда формация Кайпаровиц была расположена недалеко от западного берега Западного внутреннего моря, большого внутреннего моря, которое разделяло Северную Америку на две части: Ларамидию на западе и Аппалачию на востоке. Местность, на которой жили динозавры, несла на себе древнюю пойму, в которой преобладали крупные каналы и торфяные болота, пруды и озера, и была окаймлена горами. Климат был влажным, и поддерживал богатый и разнообразный спектр организмов. Флора и фауна этой формация является одной из самых изученных в мире.
Космоцератопс отвлекается на проходящего рядом талоса
Талос разделял среду обитания с различными животными, например тероподами из семейства дромеозаврид, овирапторид, орнитомимидами орнитомимами, тираннозавридами тератофонеями, бронированными анкилозавридами, утконосыми гадрозавридами паразауролофами и грипозаврами, цератопсами ютацератопсами, насутоцератопсами, космоцератопсами. Также в формации Кайпаровиц обитало множество амфибий, черепах, ящериц и крокодилов, а также хрящевых рыб (акул и скатов). Было множество ранних млекопитающих, в том числе представителей групп многобугорчатых, сумчатых и насекомоядных.
Сисязавр (лат. Xixiasaurus) — род троодонтид из Китая, обитавших в меловом периоде.
Название Xixiasaurus образовано от «Xixia» (по системе пиньинь так пишется название уезда Сися в Китае) и от греч. σαῦρος «ящерица», что дословно может означать «ящерица из уезда Сися».
На основании окаменелостей ученые отнесли данный род к семейству Troodontidae. При исследовании Xixiasaurus были выявлены несколько признаков характерных для троодонтидов: более высокое количество зубов, чем у большинства других тероподов, в том числе и минирапторов, таких как дромеозавриды; сужение между зубной коронкой и корнем; прорезывание зубов около рострального кончика нижней челюсти и присутствие отличного углубления для нервно-сосудистых отверстий. Тем не менее, Xixiasaurus отличается от большинства представителей семейства, но похож на Byronosaurus jaffei, Mei long, Urbacodon itemirensis и Anchiornis huxleyi в прорезывании зубов, которое является необычным для тероподов, у которых никаких зазубренности не найдены ни на предшествующих, ни на следующих этапах эволюции. Однако это отличие зубов также присутствует у некоторых других представителей, такие как альваресзаврид, Pelecanimimus polyodon и Spinosaurus aegyptiacus имеющие отличительные особенности зубов. Это, очевидно, указывает, что зазубренности были потеряны у нескольких эволюционных ветвей тероподов.
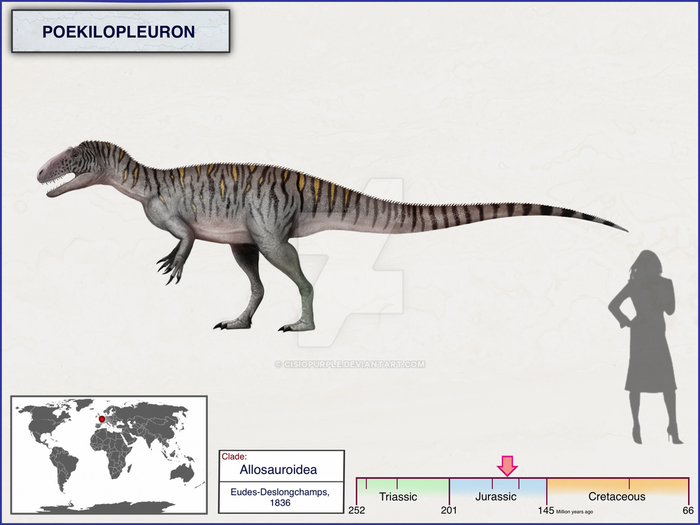
Пойкилоплеврон (лат. Poekilopleuron, что означает «разнообразные рёбра») — род тероподов-динозавров, который жил в средний юрский период (батский ярус), примерно 168–166 миллионов лет назад. Типовой вид — P. bucklandii, названный в честь Уильяма Бакленда, и многие младшие синонимы этого вида также были выделены. В настоящее время известно мало материала, так как голотип был уничтожен во время Второй мировой войны, хотя многие слепки материала всё ещё существуют.
Название Poekilopleuron происходит от греческого др.-греч. ποικίλος — «разнообразный» и πλευρόν — «ребро», из-за трёх типов рёбер. Видовое название Bucklandii дано в честь Уильяма Бакленда.
Окаменелости пойкилоплеврона происходят из среднего бата Нормандии и были описанны Эд-Делонгшаном в 1838 году, через 13 лет после мегалозавра из среднего бата Стоунзфилда в Оксфордшире. Эд-Делонгшан придал особое значение названию «Bucklandii» к его новому роду, чтобы в случае синонимии двух родов, видовое название подавляло только родовое. На основе материала голотипа, который в настоящее время уничтожен. В 1837 году Эд-Делонгшан опубликовал более подробный отчет об этом открытии в монографии, которая также была включена в следующем году в шестой том "Воспоминаний о нормандском обществе Линнея".
Слепок гастролий голотипа, Национальный музей естественной истории
Голотип, хранившийся в Музее факультета естественных наук в Кане, был уничтожен во время Второй Мировой войны, включал гастралии, фаланги, левую переднюю конечность, хвостовые позвонки, шевроны, рёбра и заднюю конечность. Из всего материала сохранилось немногое, хотя гастролиты, фаланги и передние конечности были отлиты и теперь представляют собой пластотип с отливками в Национальном музее естественной истории (образец MNHN 1897-2) и Йельском музее Пибоди (образец YPM 4938). Оригинальный материал, обнаруженный в слое Канского известняка в Нормандии, Франция, можно отнести к среднему батскому ярусу, примерно 167.7 – 166 миллионов лет назад.
В той же публикации 1836 года Эйдс-Делоншан также назвал типовой вид P. bucklandii. Эд-Деслонгшамп отметил сходство с некоторыми материалами из Megalosaurus bucklandii и P. bucklandii и выбрал видовое название bucklandii для поэкилоплеврона, так что, если бы два рода были синонимами, было бы исключено только название рода.
Кау (2024) предположил, что современный дюбрейлозавр, первоначально отнесённый к пойкилоплеврону, может представлять собой младший синоним, а различия в строении скелета объясняются позиционной и индивидуальной вариативностью, как у аллозавра. Таким образом, он представляет собой менее зрелую особь пойкилоплеврона.
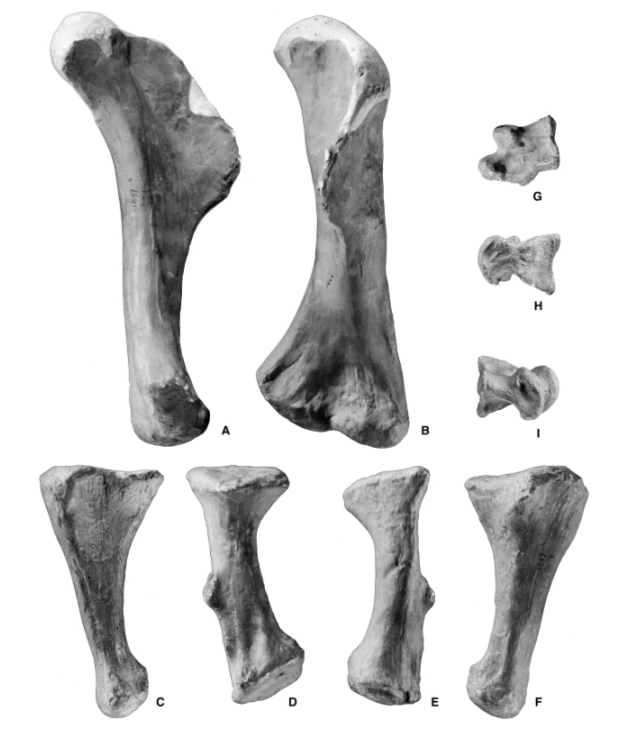
Самой отличительной особенностью пойкилоплеврона были его передние конечности. Их длина, около 60 см, что было уникально. В отличие от более поздних теропод, чьи передние конечности имели тенденцию к уменьшению в длину пропорционально размеру тела, у пойкилоплеврона были длинные, как следствие, мощные передние лапы. Длина в основном заключалась в удлиненной, но мощной мускулистой плечевой кости. Предплечья были заметно более короткими и крепкими, что характерно для немного более позднего и значительно более крупного американского родича торвозавра. Уникальной особенностью является отсутствие отростка на локтевой кости.
Кости передних конечностей
Пойкилоплеврона также сохранил редкий и полный набор гастралий: четырнадцать пар брюшных ребер, которые поддерживали тело.
Пойкилоплеврон имеет сложную историю классификации, поскольку его исходный материал утерян, и известно лишь несколько слепков. Он неоднократно переименовывался в разные виды и роды, большинство из которых теперь являются его младшими синонимами.
Эуд-Делоншан считал, что вполне возможно доказать, что образец принадлежит к ранее названному виду; если это так, то нужно будет лишь изменить родовое название. После 1879 года его часто относили пойкилоплеврона к мегалозавру. Однако выбор Эд-Делонгшана вызвал проблемы, когда Фридрих фон Хюне в 1923 году пришёл к выводу, что это был мегалозавр, но отдельный вид в рамках этого рода. Поскольку оба вида носили один и тот же видовой эпитет «bucklandii», их больше нельзя было различать. Поэтому фон Хюне переименовал вид в «Megalosaurus poekilopleuron».
Другая проблема была вызвана тем, что название было лишь частично латинизировано. На правильном греческом языке оно должно было звучать как «Poikilopleuron», на латыни — «Poecilopleurum». Это побудило более поздних авторов улучшить написание, что привело к появлению таких вариантов, как Poecilopleuron и Poikilopleuron (которые использовались до 2006 года). Однако первоначальное название имеет приоритет и является правильным.
В роде было названо ещё пять видов. В 1869 году Эдвард Дринкер Коуп переименовал «Laelaps gallicus» в «Poekilopleuron gallicum». В 1870 году Джозеф Лейди назвал «Poicilopleuron» valens на основе неполного хвостового позвонка, который, вероятно, принадлежал аллозавру. В 1876 году Ричард Оуэн назвал «Poikilopleuron pusillus», в 1879 году Коуп переименовал его в «Poekilopleuron minor»; в 1887 году Гарри Говиер Сили выделил его в отдельный род: аристозух. В 1883 году Киприянов создал «Poekilopleuron schmidti», видовое название которого дано в честь Фридриха Шмидта, на основе нескольких неопределённых рёбер, возможно ихтиозавра и плюсневой кости завропода. Эта химера является nomen dubium. Гораздо более поздний вид — «Poekilopleuron» valesdunensis, описанный Ронаном Алленом в 2002 году. В 2005 году он был переименован в дюбрейлозавра. Фрагментарные окаменелости из поздней юры и раннего мела Франции и Англии относили к пойкилоплеврону, но это является сомнительным
Реконструция черепа дюбрейлозавра, который может быть младшим синонимом пойкилоплеврона. Автор: Илья Садыков.
Поскольку оригинальная окаменелость была уничтожена, а других останков с тех пор не находили, а также из-за смены названия, вокруг его классификации возникло много споров, которые не могут быть разрешены. Традиционно его относили к мегалозавридам, но некоторые недавние анализы показали, что он относится к синрапторидам (ныне метриакантозавриды); другие результаты показали, что он является представителем мегалозавроид, занимает базальное положение или относится к эустрептоспондилинам. Бенсон и др. (2010) отнесли его и лориньянозавра к синрапторидам. Более раннее исследование показало, что он был примитивным аллозавроидом вне синрапторид. Однако недавний анализ, проведённый Каррано и др. (2012), также использованный в последующих исследованиях тетанур, показал, что он был мегалозавроидом, тесно связанным с афровенатором. Однако авторы этого исследования утверждают, что такая позиция сомнительна, поскольку «с помощью одного дополнительного шага его можно отнести к пятницкизавридам, к другим представителям мегалозаврид или к нескольким возможным представителям аллозавров».
В недавних исследованиях пойкилоплеврон был отнесён к разным группам в аллозавроидах. Кау (2024) отнёс его к метриакантозавридам, а Раугут и его коллеги отнесли его к кархародонтозаврам в 2019 и 2024 годах соответственно.
Материал, относящийся к пойкилоплеврону, включает два хвостовых позвонка, у которых шеврон одного позвонка прирос к центральной части следующего позвонка в результате развития экзостоза. Две фаланги также сохранили патологии. На одной вероятной фаланге видны три низких, неправильных выступа, похожих на экзостозы. На второй вероятной фаланге кисти имеется низкий округлый выступ, напоминающий мозоль. Ральф Молнар счёл наличие трёх патологий у одного индивида «примечательным». Образцы к сожалению могут быть исследованы для определения этиологии патологий из-за их уничтожения во время бомбардировки британцами в 1944 году.
Ихтиозавры являются наиболее адаптированными к водному образу жизни пресмыкающимися. Аналогичной степени приспособленности смогла достичь только одна группа млекопитающих — китообразные. Уникальные характеристики, связанные с водным образом жизни, наблюдаются уже у древнейших известных ихтиозавров из нижнего триаса (шпатский подъярус оленёкского яруса). К таким приспособлениям относятся удлинённая морда, очень большие глаза, преобразованные в плавники конечности и сильно вогнутые позвонки.
Скелет Ихтиозавра
Ключевой эволюционной особенностью ихтиозавров является живорождение, наиболее ранние документальные свидетельства существования которого относятся к анизийскому ярусу среднего триаса. Имели эвриапсидное строение черепа, произошли от диапсидных предков. Зубы сменялись неоднократно в течение жизни. Конечности использовались для поддержания равновесия и контроля направления. У хвоста было две лопасти, нижняя из которых поддерживалась позвоночным столбом. У типичного ихтиозавра были очень большие глаза, защищённые костяным кольцом, говорящим о том, что охотились они ночью. В связи с этим некоторые виды обладали огромными глазами (до 20 см в диаметре). Дополнительно, по-видимому, имелись какие-то кожные рецепторы, подобные боковой линии, на что указывают следы нервов и сосудов на костях черепа. Кожа, лишённая чешуи, для лучшего скольжения в воде была, возможно, покрыта слизью. Хорошо приспособлены к движению с высокой скоростью, как современный тунец. По крайней мере некоторые ихтиозавры предположительно были хорошими глубоководными ныряльщиками, как современные киты. Наиболее вероятная окраска — тёмный верх и светлый низ с синеватым оттенком. Самый крупный вид, описанный по обнаруженным окаменелостям — шонизавр из позднего триаса полярной Канады. В 2003 году описан скелет длиной около 23 метров, но в среднем длина составляла 2—4 метра.
У некоторых ранних ихтиозавров были зубы, с помощью которых они питались моллюсками — аммонитами, наутилоидеями и кальмарами. Очень похоже на то, что они также питались рыбой, а у части более крупных видов были тяжёлые челюсти и зубы, которые показывают, что они питались более мелкими рептилиями.
Анализ ископаемых остатков стеноптеригия (Stenopterygius), найденного в Германии в шахтах Хольцмадена, датированного возрастом 180 млн лет назад и сохранившего небольшие фрагменты мягких тканей, показал, что внутренние слои кожи переходили в изолирующий жировой слой как у теплокровных животных.
Некоторые ихтиозавры были очень крупными. В 2018 в Англии были найдены фрагменты нижней челюсти ихтиозавра, которые указывали на то, что он имел в длину от 20 до 25 метров, его затем описали как вид Ichthyotitan severnensis, а Ophthalmosaurus icenicus весил около тонны[9].
Типовой вид этой группы был описан в 1821 году как Ichthyosaurus communis De la Beche & Conybeare в составе рода ихтиозавров из нового семейства Ichthyosauridae De la Beche & Conybeare, 1821.
Геохронология Ихтиозавра
Ихтиозавры существовали в течение почти всего мезозоя 250—94 млн лет назад, наибольшего расцвета достигли в юрский период, пока в меловой период их не сменили плезиозавры. В мелу количество видов ихтиозавров резко снизилось, лишь один род — платиптеригиусы (Platypterygius) — дожил до начала позднего мела (единственный описанный представитель меловой группы). Предполагается, что глобальное потепление, произошедшее в середине мелового периода, повлекло за собой обеднение кислородом океанических вод, что привело к «аноксидной катастрофе» и вымиранию не приспособившихся к изменениям климата ихтиозавров.